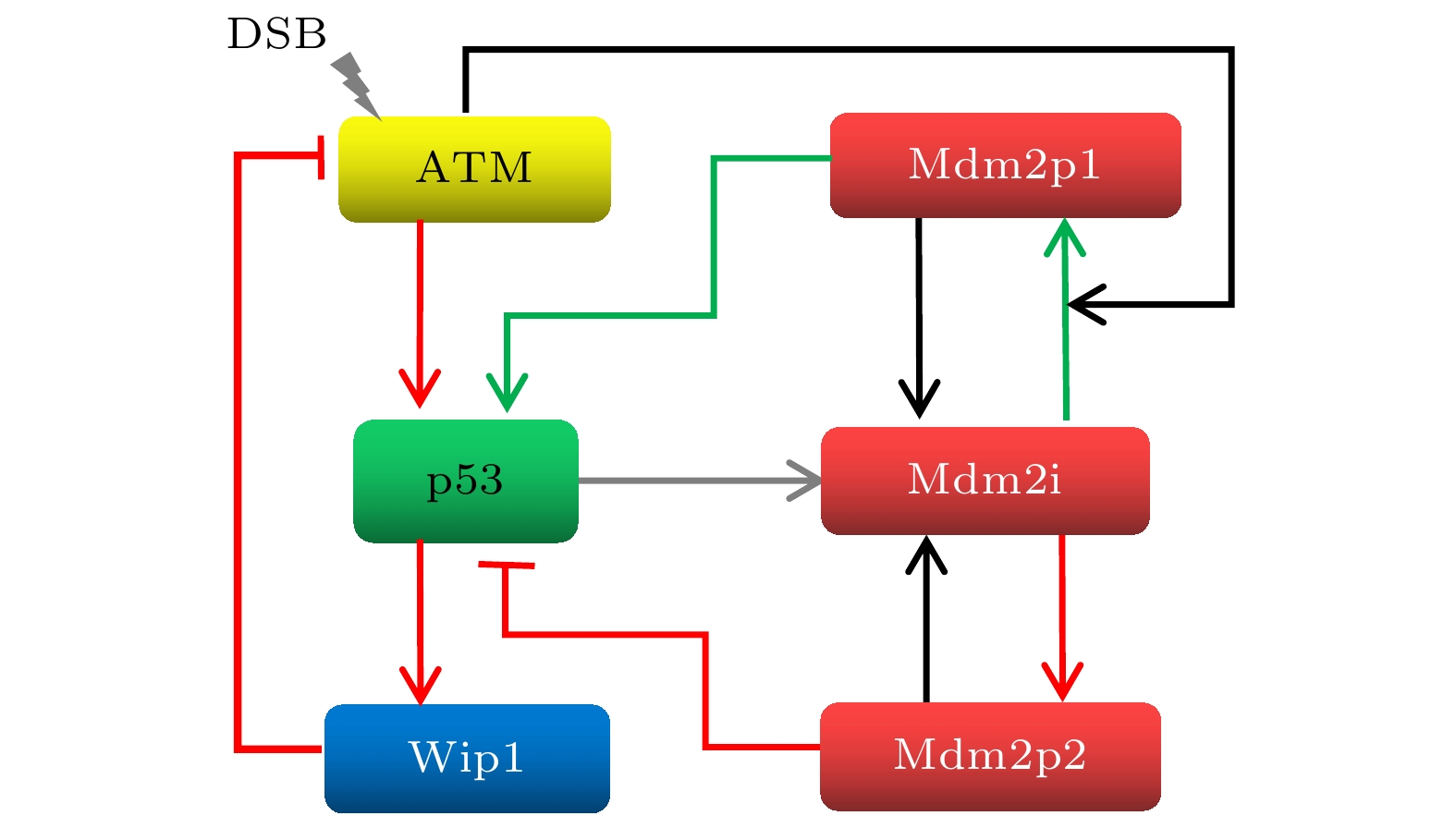
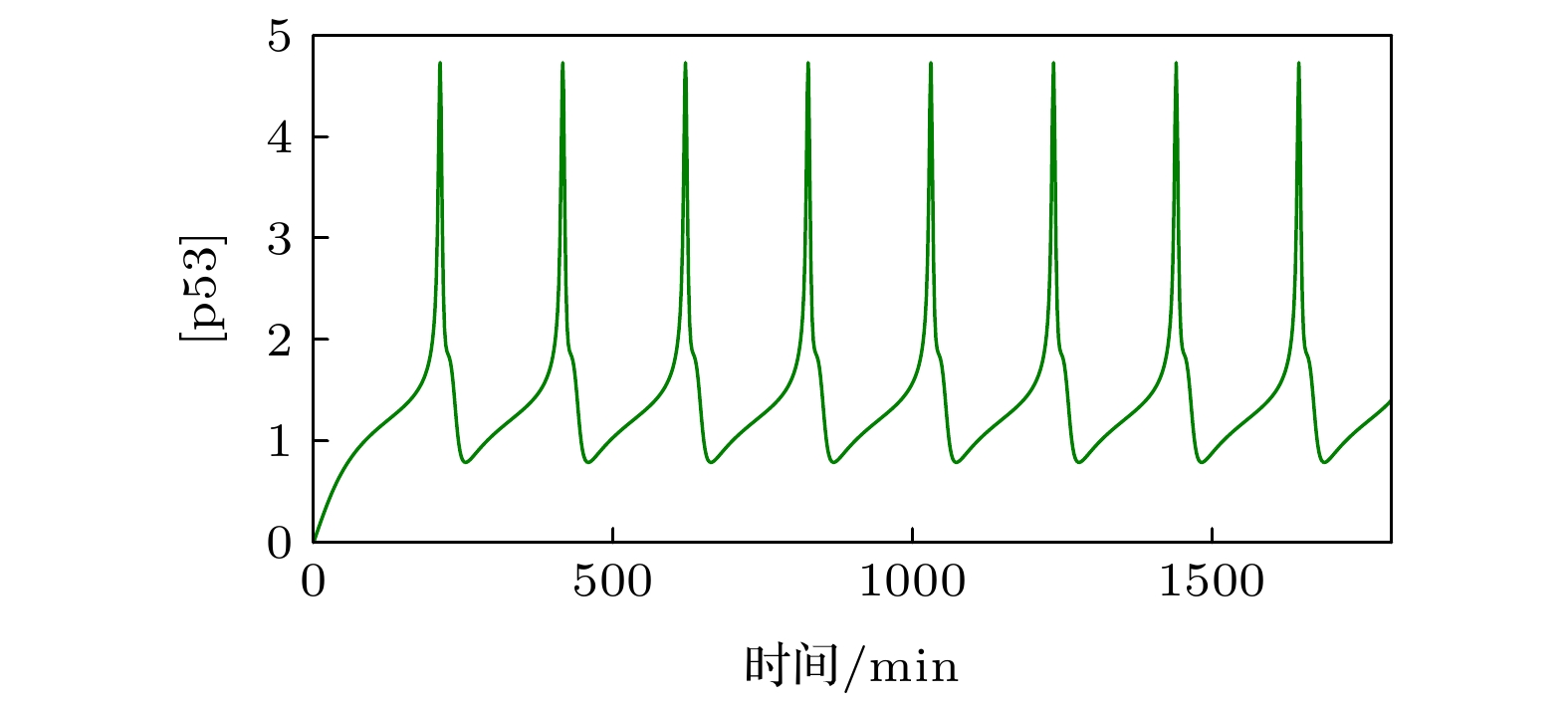
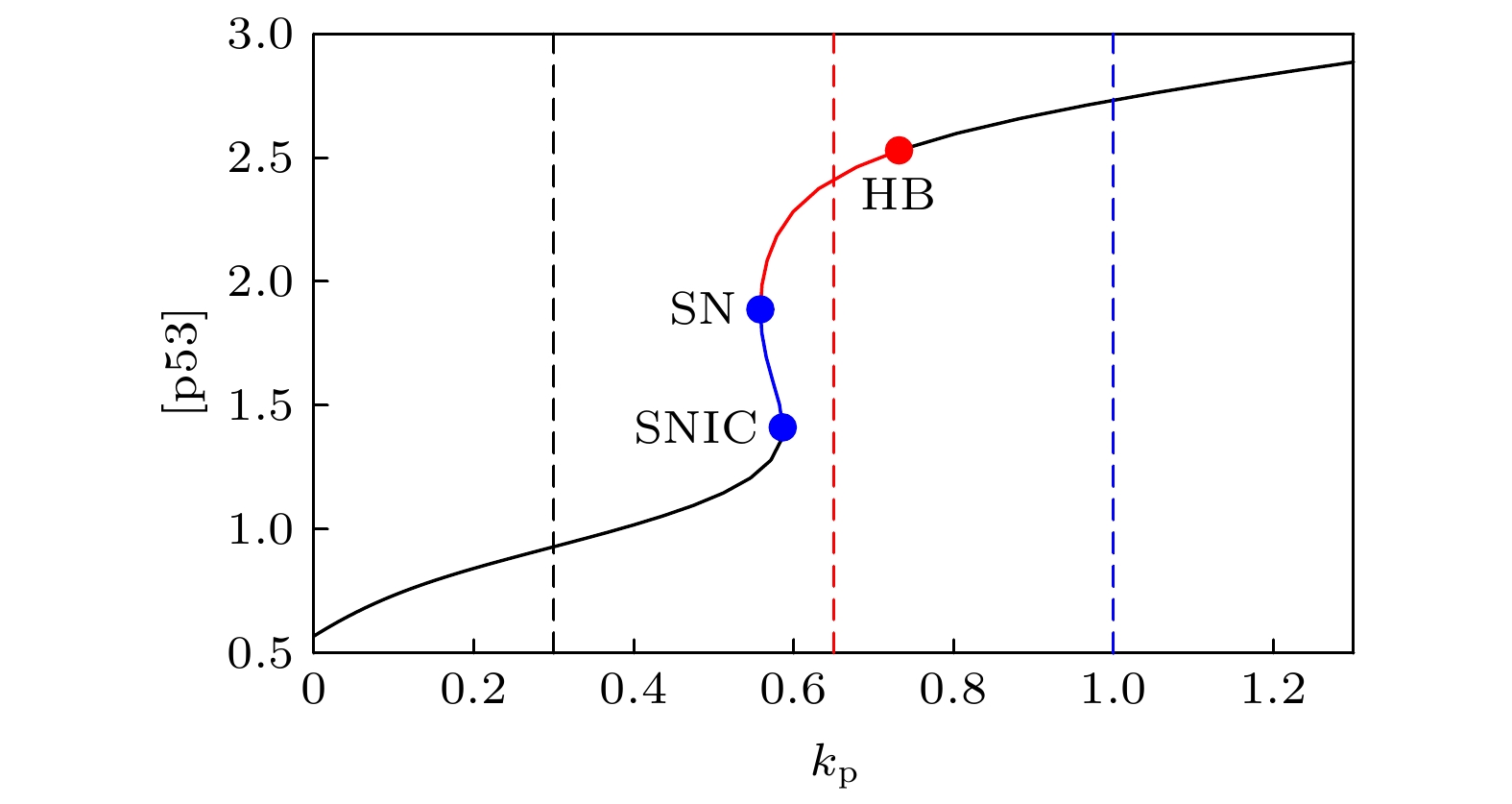
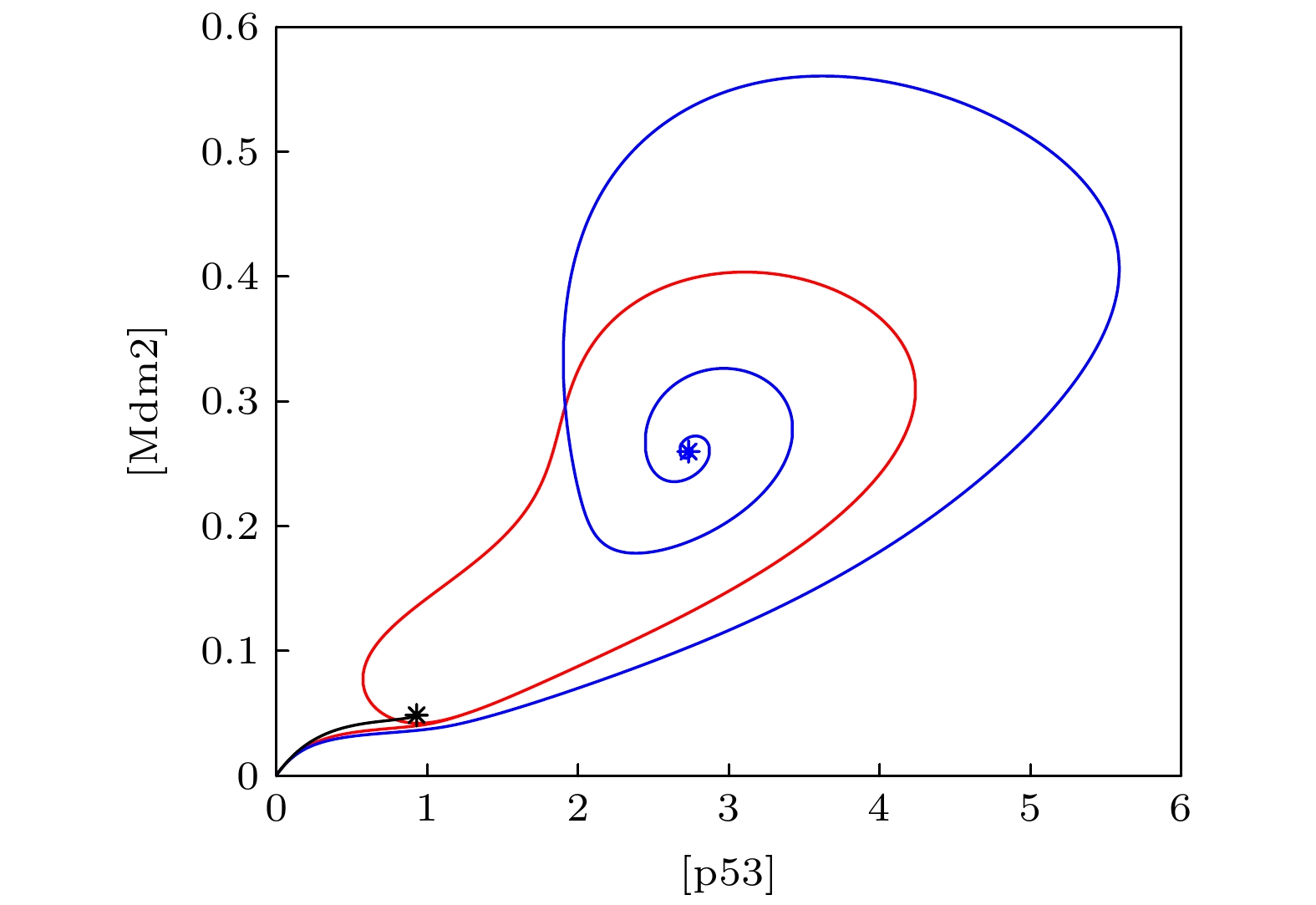
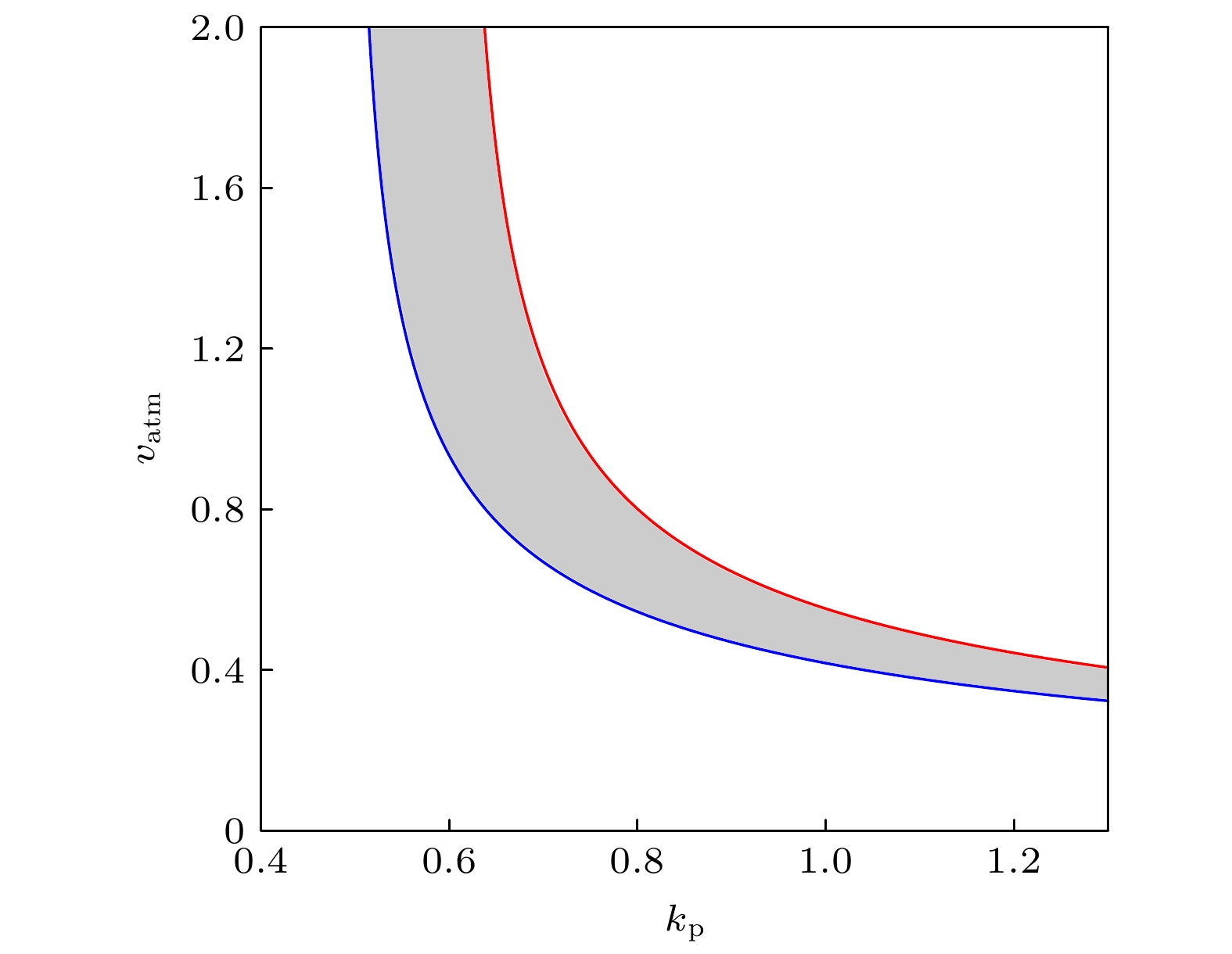
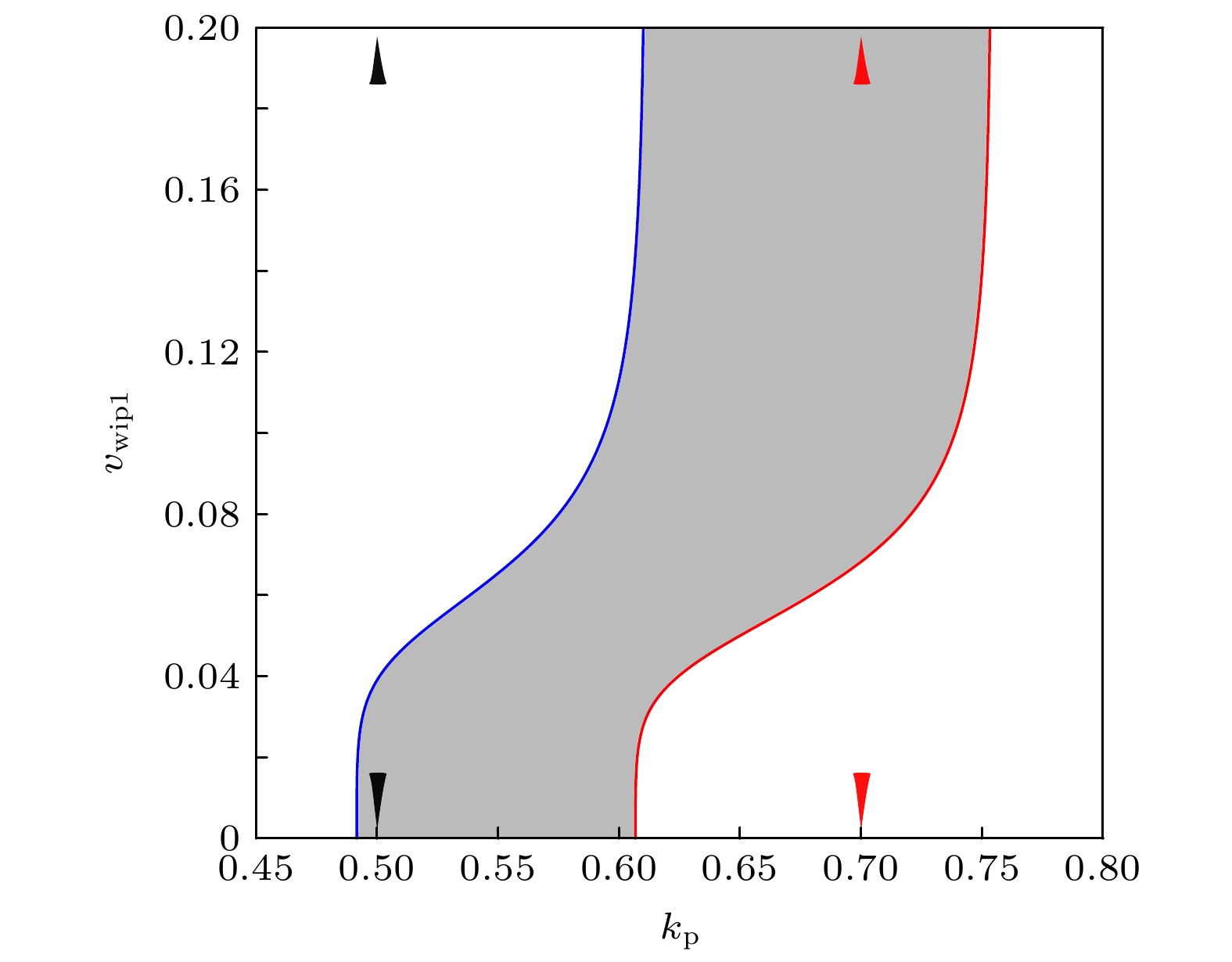
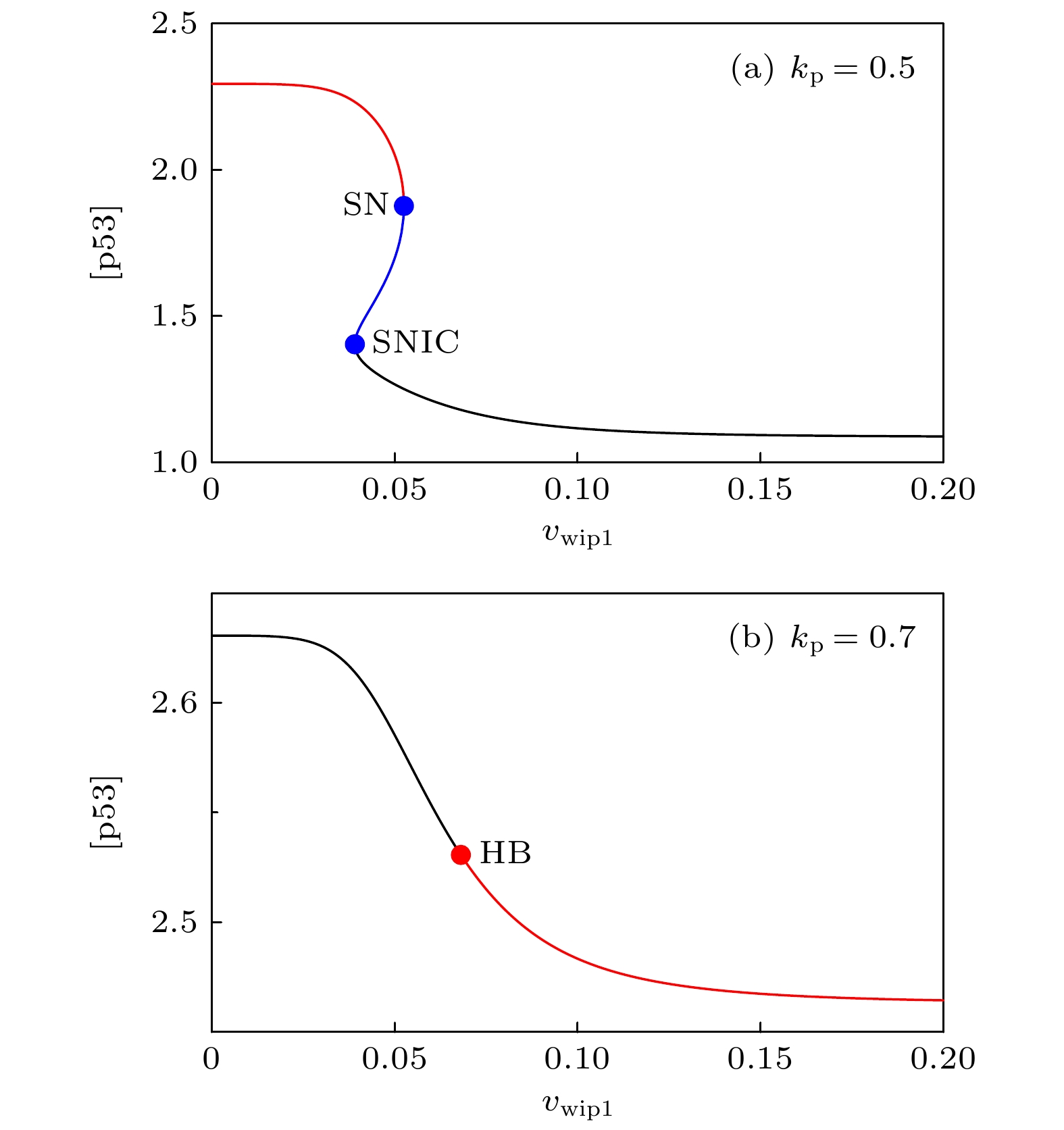
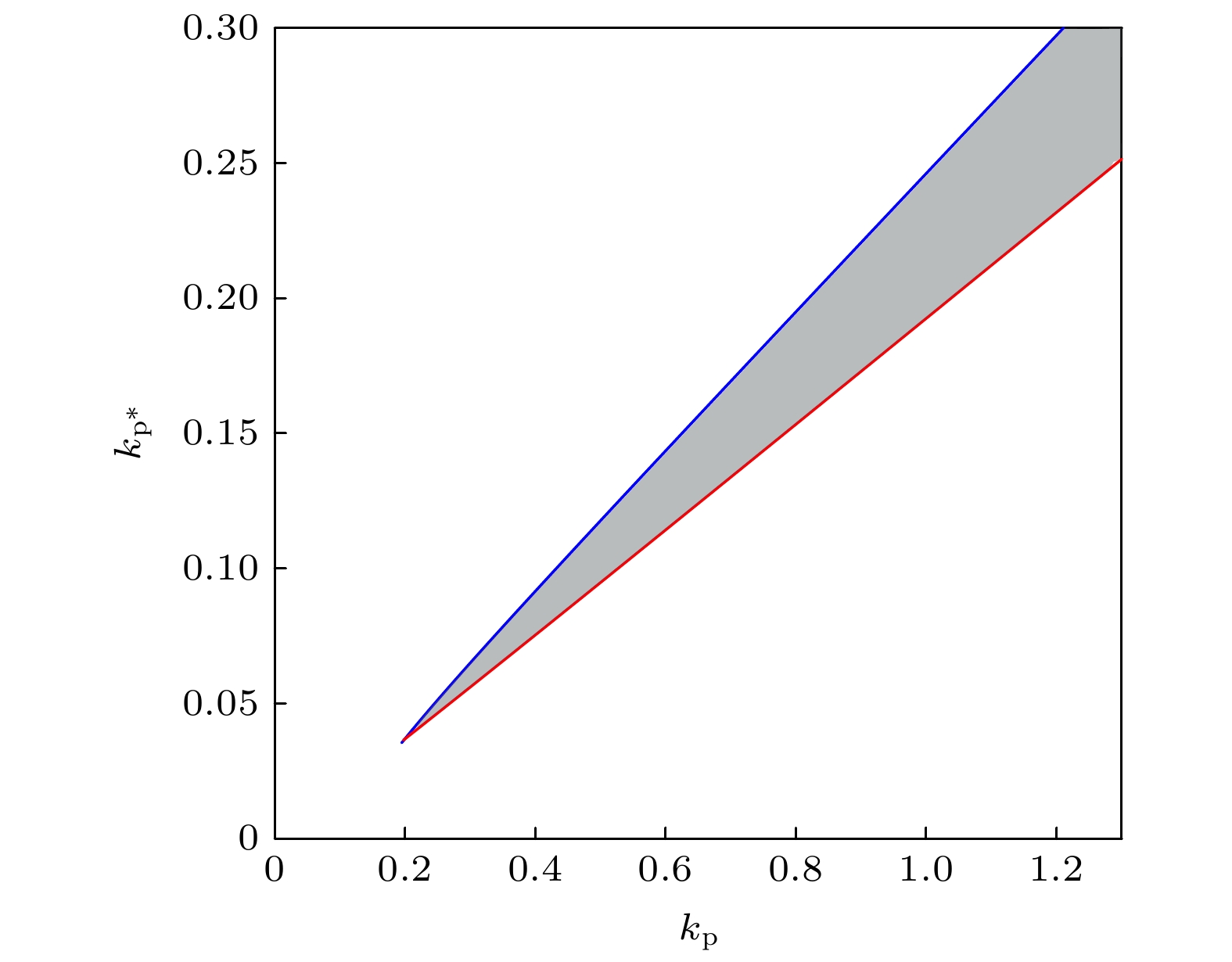
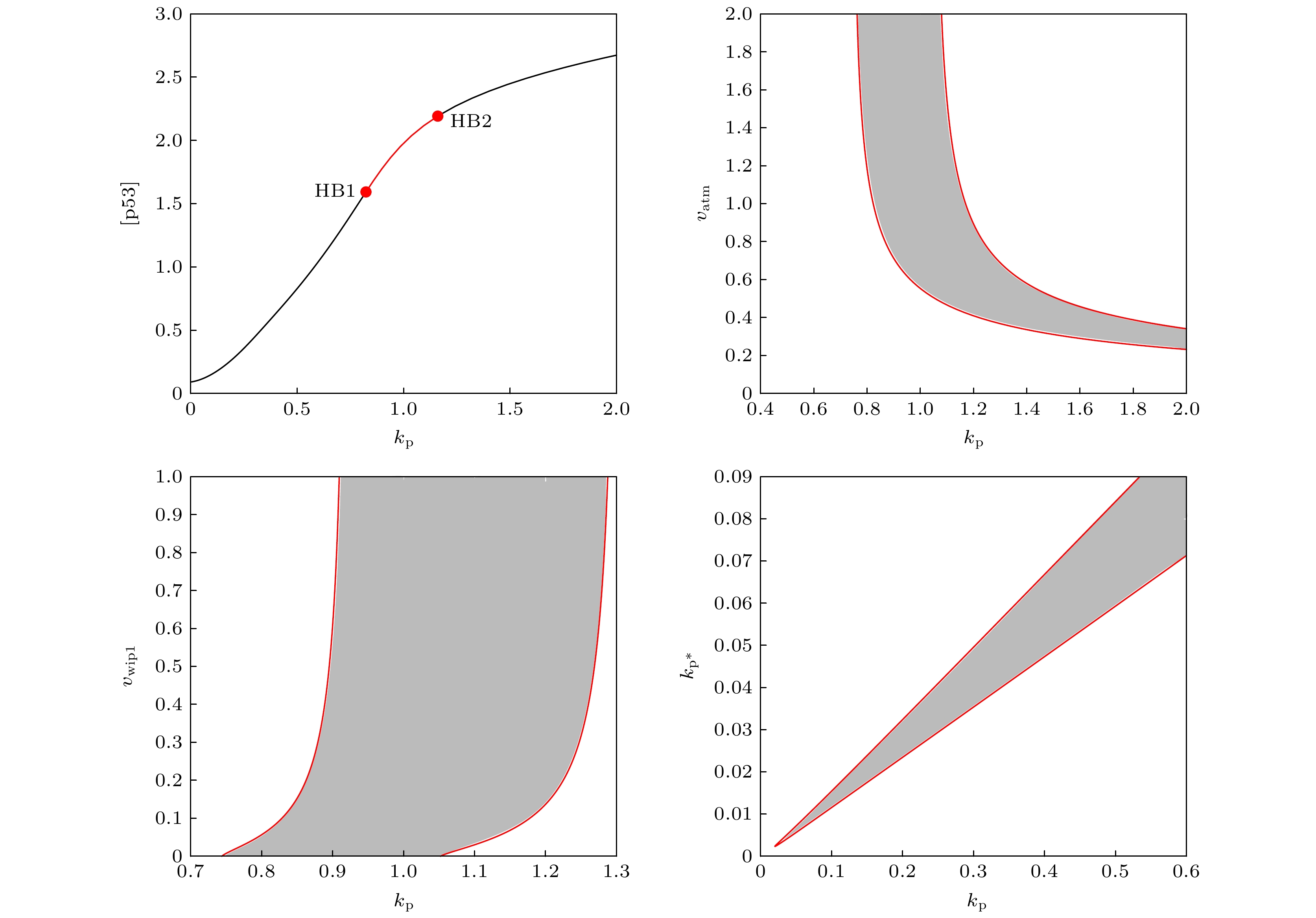
The transcription factor p53 is the core of the cellular stress network, which controls cell fate decisions on genotoxic stress in a dynamics response manner. Mdm2 is an E3 ubiquitin ligase, which play a dual role in p53 regulation, that is, on the one hand, Mdm2 destroys the stability of p53 via ubiquitin labeling, and on the other hand, Mdm2 increase the efficiency of p53 production through binding p53 messenger RNA. A number of models indicate that Mdm2's inhibitory function on p53 plays a constructive role in the p53-Mdm2 vibrator, the research of how Mdm2's promotion function on p53 regulates the dynamics of this gene network, however, is still lacking. To this end, this paper uses a mathematical model to comprehensively explore the influence of the Mdm2 up-regulation p53 pathway on p53 dynamics. We adopt the ordinary differential equations (ODE) model and the bifurcation analysis method to investigate the multiple role of Mdm2 on p53 gene networks dynamics control. The results show that the phosphorylation of Mdm2 at Ser395, which forming a positive feedback loop (PFL) with p53, is essential for the oscillation of this gene network and may be the reason why there is a bimodal switch of p53 kinetics under etoposide stimulation; the previously reported phosphatase Wip1 is required for the oscillation of p53, which may only occur in situations where the intensity of the PFL mediated by phosphorylated Mdm2 is high, and we predict that Wip1 will inhibit the p53 oscillation in some weak PFL cases; Finally, our model also proves that Mdm2 promotes the deactivation and ubiquitination degradation of p53 is a key factor in the occurrence of repeated vibration dynamics of p53. The main conclusions of this paper are exhibited in some two-dimensional bifurcation diagrams. In order to confirm these bifurcation phenomena, we discuss the case where the Hill coefficient describing the degree of nonlinearity is small. As a result, there is no obvious shape change of the bifurcation curves surrounding the oscillation region. The robustness of these conclusions is verified again. We believe that our results can play a guiding role in the future related experiments of p53 kinetics.












 DownLoad:
CSV
DownLoad:
CSV


 DownLoad:
DownLoad:







